Preliminary Assessment of the Alaska Arctic (pdf)Andy Whitehouse1 and Stephani Zador2 1Joint Institute for the Study of the Atmosphere and Ocean, University of Washington 2Resource Ecology and Fisheries Management Division, Alaska Fisheries Science Center, National Marine Fisheries Service, NOAA This preliminary assessment of the Arctic was not updated this year. We include it here as a reference for the study area and indicators that have been suggested for the development of a future full Arctic Assessment and Report Card. Defining the Alaska Arctic assessment areaIn 2012 preliminary assessment of the Alaska Arctic, we proposed the inclusion of the northern Bering Sea (>approx. 60°N) within the Alaska Arctic assessment area. The Alaska Arctic assessment area would then include the entire Arctic management area (NPFMC, 2009) and the northern Bering Sea (Figure 15). This suggestion was made in recognition of the growing body of scientific literature that indicates the northern Bering Sea is biologically and physically distinct from the southeastern Bering Sea (Grebmeier et al., 2006; Mueter and Litzow, 2008; Sigler et al., 2011; Stabeno et al., 2012; Stevenson and Lauth, 2012). The northern Bering Sea is not presently part of the assessed area in the eastern Bering Sea. Thus including the northern Bering Sea within the proposed Arctic area would create a continuum of assessed large marine ecosystems (LMEs) throughout Alaska. In the time since our preliminary assessment was published, the Arctic Council, an international forum of Arctic governments and indigenous communities (http://www.arctic-council.org), has published a revision to their boundaries for LMEs of the Arctic Area (PAME, 2013). In their revision they moved the southern boundary of the Chukchi LME further south into the northern Bering Sea. Previously their boundary was at the Bering Strait (˜66°N) but is now located south of St. Lawrence Island at 61.°N. Similarly, the rationalefor this revision was in recognition of the combined biological and physical properties linking the northern Bering Sea to the Chukchi Sea. As this Arctic section of the Ecosystem Considerations report progresses we will likely specify 61.°N as the southern boundary of the Alaska Arctic assessed area, coincident with the LME boundary revisions made by the Arctic Council.
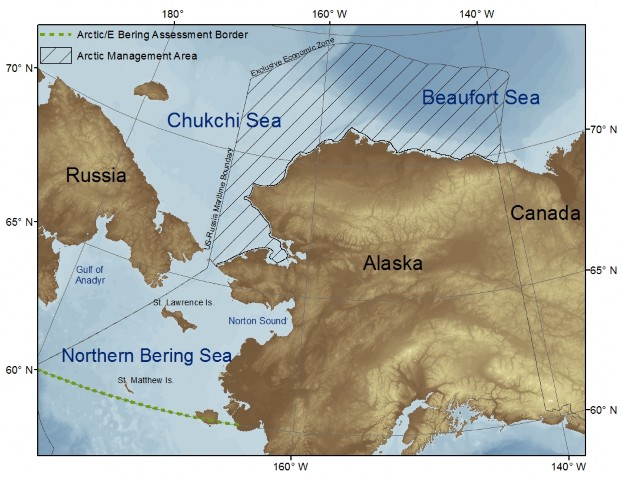
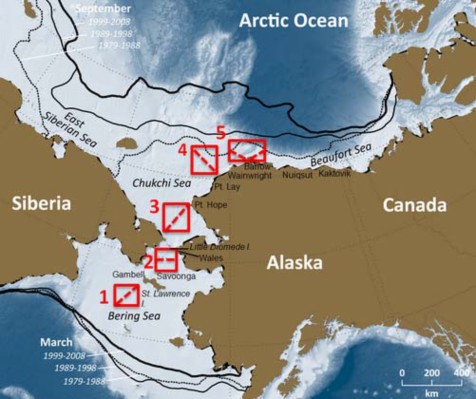
Figure 15: The proposed Arctic assessment area in Alaska, encompassing the northern Bering Sea, Chukchi Sea, and Beaufort Sea, within US territorial waters. The existing Arctic management area is filled with hatched lines. General ecosystem informationMost of the Alaska Arctic is covered by sea ice for some portion of the year and the seasonal presence and dynamics of sea ice has a strong influence on ecosystem structure and function. During years of low ice coverage, the most southerly portions of the northern Bering Sea may only be covered by sea-ice for a few weeks or not at all. The Chukchi and Beaufort seas are covered by sea ice for about 6 to 8 months of the year. During years of heavy summer ice coverage, portions of the northern Chukchi and Beaufort seas may retain their ice coverage throughout the year. However, Arctic sea ice cover has declined over recent decades, with the seven lowest annual sea ice minima over the satellite record (1979-present) occurring in the last 7 years, 2007-2013 (Comiso, 2012; Stroeve et al., 2012)(http://nsidc.org). A recent reconstruction of Arctic sea ice cover over the last 1,450 years has indicated that the observed declines in sea ice starting in the 1990's are the lowest over this time period, and fall outside the range of variability in previous observations (Kinnard et al., 2011). Regionally, some of the most pronounced declines of September ice extent in recent decades have been observed in the Chukchi and Beaufort seas (Meier et al., 2007). The persistence of sea ice during the summer season has implications for the primary productivity regimes in these northern systems. Primary production during winter is limited by ice coverage and shortened day length, including periods of arctic night in the Chukchi and Beaufort seas. Phytoplankton growth begins in late winter with the return of daylight and an ice algae bloom that continues until the onset of ice melt (Cota, 1985; Cota and Smith, 1991)). At a time when food may be limited, the ice algae bloom provides early season forage for ice-associated invertebrates, which in turn are preyed upon by Arctic cod Boreogadus saida ) (Bradstreet and Cross, 1982; Legendre et al., 1992; Gradinger and Bluhm, 2004). In seasonally ice covered areas, ice algae may contribute less than 5% to total annual primary production (water column and sea ice), while at the northern margins of the Chukchi and Beaufort seas, which may experience year-round ice coverage, ice algae can account for more than 50% of the annual primary production budget (Gosselin et al., 1997). Additionally, recent work in the northern Chukchi Sea has indicated that under-ice phytoplankton blooms, which had previously been unaccounted for, may contribute substantially to total primary production (Arrigo et al., 2012). Current estimates of primary production over Arctic continental shelves that do not take these under-ice blooms into account may be several times too low (Arrigo et al., 2012). The breaking-up and melting of sea ice in spring strengthens water column stratification, and when combined with increasing day-length, induces an ice edge phytoplankton bloom that follows the retreating ice edge northward (McRoy and Goering, 1974; Niebauer et al., 1981; Sakshaug, 2004). Seasonal ice coverage cools the entire water column over the shallow shelves of the northern Bering and Chukchi seas to temperatures below 0°C. These cold temperatures limit the northern distribution of sub-Arctic populations of groundfish, such as walleye pollock and Pacific cod (Osuga and Feeney, 1978; Wyllie-Echeverria and Wooster, 1998; Mueter and Litzow, 2008; Stevenson and Lauth, 2012), and may constrain their growth (Pauly, 1980). During summer much of the zooplankton community occupying the northern Bering and Chukchi seas are of Pacific origin, and are advected into these Arctic waters through Bering Strait (Springer et al., 1989; Hopcroft et al., 2010; Matsuno et al., 2011). Here, the cold water temperatures may limit zooplankton growth and their grazing efficiency of phytoplankton (Coyle and Pinchuk, 2002; Matsuno et al., 2011). Cold-adapted Arctic zooplankton species are more prevalent in the northern portions of the Chukchi Sea, near the continental slope and canyons (Lane et al., 2008). In years of low ice coverage, an overall northward distribution shift in southern extent of Arctic species and the northern extent of Pacific species hasbeen observed (Matsuno et al., 2011). Additionally, an increase in total zooplankton abundance and biomass has also been observed in years of low ice coverage, and this has been in part attributed to an increased influx of larger zooplankton species of Pacific origin and temperature effects on their growth (Matsuno et al., 2011). The annual dynamics of sea ice also affects the distribution of marine mammals. Pacific walrus and ice seals utilize sea ice in the Bering Sea during winter to haulout, breed, and whelp. Ringed seals are present throughout the Alaska Arctic during winter and maintain breathing holes in the ice to keep access to the water (Lowry et al., 1980; Kelly, 1988). Ringed seals also construct resting lairs over breathing holes and beneath the snow cover, which provide protection from the elements and predators, and are used to raise pups (Burns, 1970; Smith et al., 1991; Kelly et al., 2010). Pinnipeds may also use sea ice as a form of transportation during ice retreat and as a platform to rest between foraging excursions. Polar bears utilize sea ice as platform to hunt from throughout the year. Pregnant female polar bears may also excavate maternity dens on sea ice in the fall, where they will give birth to cubs in winter (Lentfer and Hensel, 1980; Amstrup and Gardner, 1994; Fischbach et al., 2007). Belugas and bowhead whales spend the winter along the ice edge in the northern Bering Sea, and in the spring they follow regularly recurring leads and fractures in the ice that roughly follow the Alaska coast during migration toward their summering grounds in the Beaufort Sea (Frost et al., 1983; Ljungblad et al., 1986; Moore and Reeves, 1993; Quakenbush et al., 2010). Belugas also forage near the ice edge and in more dense ice coverage among leads and polynyas in both the Beaufort and Chukchi seas (Richard et al., 2001; Suydam, 2009). Seabirds may also concentrate near the ice-edge (Divoky, 1976; Bradstreet and Cross, 1982; Hunt, 1991), preying on ice-associated invertebrates and Arctic cod (Bradstreet and Cross, 1982). Marine mammals have been important subsistence resources in Alaska for thousands of years and the continued subsistence harvests of marine mammals are important to the maintenance of cultural and community identities (Hovelsrud et al., 2008). The presence and dynamics of sea ice is an integral part of many subsistence harvests, including the hunting of bowhead whales (George et al., 2004), belugas (Huntington et al., 1999), Pacific walrus (Fay, 1982), and ice seals (Kenyon, 1962). Traditional knowledge of sea ice behavior, the effect of environmental conditions on sea ice stability, and how sea ice conditions relate to the seasonal presence and migratory habits of marine mammals has accumulated over time. The sharing of this knowledge helps maintain the successful and safe harvest of marine mammals (Huntington et al., 1999; George et al., 2004; Noongwook et al., 2007). The net flow of water through the northern Bering and Chukchi seas is northward (Coachman et al., 1975; Walsh et al., 1989; Woodgate et al., 2005). High levels of primary production in the northern Bering and southern Chukchi seas is maintained throughout the open water season by nutrient rich water advected from the Bering Sea continental slope and the Gulf of Anadyr (Springer and McRoy, 1993; Springer et al., 1996). During the open water season, primary production in the northern Chukchi Sea is focused in the vicinity of the ice edge (Wang et al., 2005) and Barrow Canyon where occasional flow reversals allow for upwelling of Arctic basin waters, which promote phytoplankton blooms (Aagaard and Roach, 1990; Hill and Cota, 2005; Woodgate et al., 2005). Primary production in the Beaufort Sea may be enhanced during summer when sea ice retreats beyond the shelf break allowing for phytoplankton blooms driven by upwelling along the shelf break (Pickart et al., 2009). The northern Bering and Chukchi seas are benthic-dominated systems. Several ecological studies carried out over the last approximately 50 years have documented the abundant community of benthic invertebrates (Sparks and Pereyra, 1966; Feder and Jewett, 1978; Stoker, 1981; Grebmeieet al., 1988; Feder et al., 1994, 2005, 2007; Bluhm et al., 2009). Here, the combination of high primary production, shallow continental shelves (< 60 m), and cold water limiting the growth and grazing of zooplankton results in high delivery of organic matter to the benthos, where it supports an abundant benthic community (Grebmeier et al., 1988; Grebmeier and McRoy, 1989; Dunton et al., 2005; Lovvorn et al., 2005). The prominent benthos supports a community of benthic-foraging specialists, including gray whale (Highsmith and Coyle, 1992), Pacific walrus (Fay, 1982), bearded seals (Lowry et al., 1980), and diving ducks (eiders) (Lovvorn et al., 2003). Species of commercial interestSnow crabs are the basis of an economically important fishery in the eastern Bering Sea (NPFMC, 2011) and are a species of potential commercial importance in the Alaska Arctic (NPFMC, 2009). Snow crab are a dominant benthic species in the Chukchi and Beaufort seas. However, they are seldom found to grow to a commercially viable size, which is >78 mm carapace width (CW) (Frost et al., 1983; Paul et al., 1997; Fair and Nelson, 1999; Bluhm et al., 2009). More recently, a trawl survey of the western Beaufort Sea in August 2008 (Rand and Logerwell, 2011) documented the first records of snow crab in the Beaufort Sea at sizes equal to, or greater than the minimum legal size in the eastern Bering Sea, finding males as large as 119 mm CW. Studies of snow crab reproduction biology have observed some flexibility in the size at maturation, indicating snow crabs in these colder Arctic waters may mature at a smaller size (Somerton, 1981; Paul et al., 1997; Orensanz et al., 2007). Snow crabs are also found throughout the northern Bering Sea. Commercially important species of king crab have been sparsely encountered in the Chukchi Sea (Barber et al., 1994; Fair and Nelson, 1999; Feder et al., 2005) and were not encountered during the 2008 survey of the western Beaufort Sea (Rand and Logerwell, 2011). In the northern Bering Sea blue king crab are found near St. Matthew Island and north of St. Lawrence Island, and red king crab in Norton Sound (Lauth, 2011). The northern Bering Sea (as defined here) includes the northern half of the Alaska Dept. of Fish & Game management area for St. Matthew Island blue king crab. Following a ten year closure to rebuild the St. Matthew Island stock of blue king crab, the commercial fishery was reopened in 2009/10 (NPFMC, 2011). Red king crab presently support both, commercial and subsistence fisheries in Norton Sound (NPFMC, 2011). The fish resources of the Alaska Arctic have not been as thoroughly sampled as in other large marine ecosystems in Alaska (e.g., eastern Bering Sea, Gulf of Alaska, Aleutian Islands), but a limited number of standardized demersal trawl surveys have been conducted in the region since the mid 1970's. The northern Bering and southeastern Chukchi seas were surveyed in 1976 (Wolotira et al. 1977), the northeastern Chukchi Sea in 1990 (Barber et al., 1994, 1997), the western Beaufort Sea in 2008 (Rand and Logerwell, 2011), the northern Bering Sea again in 2010 (Lauth, 2011),and the eastern Chukchi Seat in 2012 (Arctic EIS, https://web.sfos.uaf.edu/wordpress/arcticeis/). The catch data from these trawl surveys indicate that fish sizes are generally small and demersal fish biomass is low. Though fish have not been particularly abundant in survey catches, when present they have been dominated by cods, flatfishes, sculpins, and eelpouts (Wolotira et al., 1977; Barber et al., 1997; Lauth, 2011; Rand and Logerwell, 2011). In the Chukchi and Beaufort seas, Arctic cod has been consistently identified as the most abundant fish species (Alverson and Wilimovsky, 1966; Quast, 1974; Wolotira et al., 1977; Frost et al., 1983; Barber et al., 1997; Rand and Logerwell, 2011). They occur in benthic and pelagic habitats in ice-free waters and are also found in association with sea-ice during ice covered periods (Bradstreet et al., 1986; Gradinger and Bluhm, 2004; Parker- Stetter et al., 2011). Arctic cod primarily prey on pelagic and ice-associated invertebrates and also form an important prey base for pelagic predators, including belugas, seabirds, and ice seals (Bradstreet and Cross, 1982; Frost and Lowry, 1984; Welch et al., 1992). Commercially important species of the eastern Bering Sea, such as walleye pollock and Pacific cod, have been infrequently encountered in the Chukchi and Beaufort seas (Frost et al., 1983; Barber et al., 1997; Norcross et al., 2010; Rand and Logerwell, 2011). Gaps and needs for future Arctic assessmentsThe intent of adding the Alaska Arctic to the regions assessed in the Ecosystem Considerations report is to provide information placed within a broad ecosystem context to fisheries managers that would be useful when making decisions on the authorization and management of new fisheries in the Alaska Arctic. We intend for future Arctic assessments to include indicators that directly address ecosystem-level processes and attributes that can inform fishery management advice. There is a continued need to convene Arctic experts to identify a list of indicators and corresponding time series data that best capture ecosystem components and trends that would be of value to fishery managers. Several biomass indices are presently used as indicators in assessments of the EBS, GOA, and AI. Time series data to support similar indices in the Alaska Arctic are lacking, but recent ongoing studies are accumulating data that may be of use as indicators. Several data sets that may be of future use are being collected by the Distributed Biological Observatory (DBO,http://www.arctic.noaa.gov/dbo/index.html. The DBO is a coordinated effort by international members of the Pacific Arctic Group (PAG, http://pag.arcticportal. org) ) that has begun to collect scientific observations at selected locations (transects) over a latitudinal gradient from the northern Bering Sea to the western Beaufort Sea, in an effort to track ecosystem change over time (Figure 16) ). As data accumulate, it is hoped that the sampling design of the DBO across a range of latitude will permit it to detect emergent patterns and trends. The data to be collected include oceanographic measurements (temperature, chlorophyll, etc.) and biological measurements, such as species composition, biomass, and the size and condition of selected key species (Grebmeier et al., 2010). Many of these metrics may be suitable for use as indicators in future Arctic assessments. Potential indicatorsIn the 2013 preliminary Arctic assessment we suggested a short list of potential indicators as a starting point for indicator discussion and development. in 2014 we presented an expanded list that includes the indicators suggested in the 2013 document, some of which are presently available (both climate indicators), and some additional biological indicators that may be of value, but are not presently available. The compiled list of potential indicators includes: Climate
Plankton
Fish
Seabirds
Marine mammals
Humans
|

Arctic Assessment (2015 update)