The Influence of Polychaete Worm-Tube Habitat on the Food Habits, Prey Availability, and Condition of Juvenile Northern Rock Sole in a Coastal Nursery off Kodiak Island, Alaska
From an essential fish habitat perspective, the role of small-scale biogenic habitat structures has not garnered the same level of interest that the role of more conspicuous macro-invertebrates, such as deep sea corals and sponges, has. Nonetheless, juveniles of many commercially important fishes show a strong association with small-scale structures, such as polychaete worm tubes which can form extensive mats on the seafloor. In embayments around Kodiak Island, Alaska, age-0 northern rock sole (Lepidopsetta polyxystra) are the most common member of the juvenile flatfish assemblage during the summer. Field studies reveal that peak northern rock sole densities occur at depths less than 30 m and are closely associated with seafloor characterized by small-scale biogenic structures, most notably tube-building, ampharetid polychaetes (Sabellides sibirica).
We hypothesized that juvenile northern rock sole were attracted to areas of worm-tube habitat to feed directly on the worms or associated fauna. We further hypothesized that worm-tube habitat would contain higher densities of benthic infauna (potential prey) compared to adjacent areas devoid of worms, thereby creating enhanced feeding conditions for the juvenile northern rock sole. Lastly, we hypothesized that northern rock sole condition would be higher in worm-tube habitat, compared to areas devoid of worms, as a result of more abundant food resources.
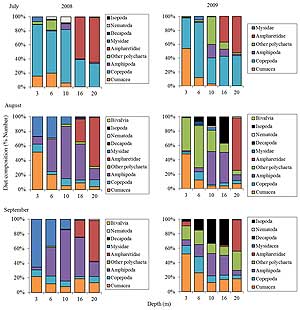
Figure 5. (Click to enlarge.) Diet composition (percentage by number) of age-0 northern rock sole collected at Pillar Creek Cove during the summer months of 2008 and 2009.
In the summer of 2008 and 2009, age-0 northern rock sole were collected for stomach content and condition analyses at the Pillar Creek Cove (PCC) study site in Monashka Bay along the northeastern coast of Kodiak Island, Alaska. Beam trawl collections were conducted along five fixed-position parallel transects oriented parallel to shore, at depths of 3-20 m. Prior video surveys at this site indicated the shallowest transect lines (3, 6, and 10 m) were positioned over mostly sand substrate, while the two deepest transects (16 and 20 m) were located over areas of sparse to moderate worm-tube density. Diet composition was described using percentage frequency of occurrence and percentage numerical importance. Condition was assessed by using log-transformed length-weight regression residuals as an indicator of somatic growth. Positive length-weight residuals indicate fish are in better condition, whereas negative residuals indicate fish are in poorer condition. Lastly, benthic grabs were made along the same transect lines in July and August 2008 to characterize the benthic fauna to determine the influence of depth and habitat (worm tubes) upon benthic faunal density and species composition.
A total of 505 age-0 northern rock sole were collected for stomach content analyses in 2008 and 2009. In each sampling period, depth and the presence of worm-tube habitat had a major influence on northern rock sole diet composition, and there were several noticeable trends (Fig. 5). In July, the northern rock sole's diet was relatively restricted and was made up primarily of harpacticoid copepods, though the importance of copepods decreased with depth. Cumaceans, mysids, and errant polychaetes were also important prey items in the shallow depths during August and September. Gammarid amphipods were generally the most important prey in the intermediate depths. Ampharetid worms were typically not consumed at depths less than 16 m, inshore of the worm-tube habitat. However, they were consistently the most important prey for northern rock sole at the deepest depth transect located in the worm-tube habitat. These results provide support for our first hypothesis by demonstrating that juvenile northern rock sole in the worm- tube habitat were feeding on the worms.
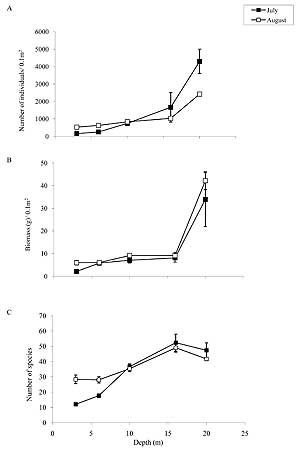
Figure 6. A- Mean number of individuals (± SE), B- Mean biomass (± SE), and C- Mean number of species of benthic infauna from Pillar Creek Cove over a range of depths during July and August 2008.
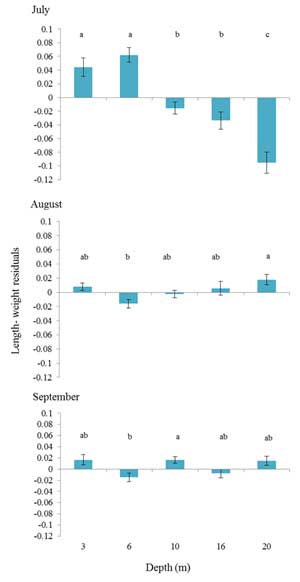
Figure 7. Mean length-weight residuals (± SE) for age-0 northern rock sole over a range of depths during July-August 2009 in Pillar Creek Cove. Letters indicate significant differences (Tukey test, P < 0.05).
As expected, depth had a pronounced influence upon both the number and biomass of benthic invertebrates. The total number of individual organisms increased with depth, and in each month the number of individuals increased most dramatically at the greatest depth sampled (Fig. 6). Correspondingly, the total invertebrate biomass also increased significantly beyond 16-m depth. Additionally, the number of species increased with depth and peaked at 16 m in each month. Each of these measures increased most dramatically with the presence of the ampharetid worm tube. This suggests that the worms may act as a bioengineering species, creating habitat that is favorable to other species, thereby increasing overall benthic infauna biomass and density.
In general, northern rock sole body condition varied with depth, but a clear trend was only evident during July (Fig. 7). In July, the length-weight residuals decreased with depth, with the highest residuals at the two shallowest depths. In August and September, northern rock sole condition varied with depth, but no discernable pattern was evident. These findings show that depth-related effects on rock sole body condition were most significant shortly after northern rock sole settled into the coastal nurseries during July. The influence of depth and habitat (worm tubes) on northern rock sole condition diminished over the course of the summer as the juvenile northern rock sole grew and consumed a more diverse diet.
Overall, it was clear that northern rock sole associated with the worm-tube habitat fed predominantly on the ampharetid worms. Additionally, the results indicate the worm-tube habitat had a positive influence on the density and diversity of benthic infauna. These results provide support for the hypotheses that juvenile northern rock sole are attracted to worm-tube habitat to feed on the worms and that worm-tube habitat offers enhanced feeding opportunities for juvenile northern rock sole compared with areas devoid of worms.
However, higher densities of benthic infauna did not result in higher condition values for northern rock sole residing in the worm-tube habitat. In fact, an opposite pattern was observed, but only in July. It is possible that the residual index we used, which is a reflection of somatic growth, was not sensitive enough to detect fine-scale spatial differences in condition during the latter months. A more sensitive biochemical condition index, such as RNA:DNA ratios, or more direct measurements of condition (e.g., bomb calorimetry) may be more suitable in detecting short-term changes in northern rock sole condition. The observed spatial variations in condition may reflect differences in total energy intake of rock sole among the depths. Differences in energy intake would result from not only differences in prey energy content but also differences in the costs associated with finding, capturing, and handling prey among the depths. It is possible that the energetic costs of finding and capturing prey were less in the sand habitat as compared to the more complex worm-tube habitat. Furthermore, the worm-tube habitat also supports high abundances of other juvenile flatfish species and crustaceans that feed on similar prey. As a result, interspecific competition might limit food resources for juvenile northern rock sole, especially when their diet was restricted in July.
At this time, the value of the worm-tube habitat in terms of northern rock sole condition and growth remains unclear. Further studies are needed to investigate the factors influencing juvenile northern rock sole condition and growth in relation to habitat features within the nursery. Regardless, it is clear that the ampharetid worm-tube habitat altered the foraging behavior of juvenile rock sole. The worms were an important food source, though they were spatially restricted to the deeper depths. In addition, the worm-tube habitat supported higher densities of benthic infauna compared to adjacent areas devoid of worms. The worm-tube habitat could have implications for both individual northern rock sole growth and overall nursery productivity by altering both the quantity and quality of available prey resources. This research highlights the importance of understanding the mechanisms influencing fish-habitat relationships which is a fundamental principle of ecosystem-based management.