The eastern Bering Sea (EBS) shelf is a highly productive ocean region, which responds rapidly to changes in climate. Because of its economic and environmental importance and climate sensitivity, the U.S. National Science Foundation's Bering Ecosystem Study (BEST) and the North Pacific Research Board's (NPRB) Bering Sea Integrated Ecosystem Research Program (BSIERP) combined to form the Bering Sea Project to study the impacts of climate change and dynamic sea-ice cover on the ecosystem of the EBS.
As part of the Bering Sea Project, the Alaska Fisheries Science Center's National Marine Mammal Laboratory (NMML) collected cetacean sightings data to assess distribution, estimate abundance, and estimate trends in abundance of cetaceans, particularly fin whales (Balaenoptera physalus) and humpback whales (Megaptera novaeangliae), on the eastern Bering Sea shelf.
Because surveys to determine distribution and abundance in the EBS are costly, NMML teamed with the Center's Resource Assessment and Conservation Engineering (RACE) division to conduct visual surveys for cetaceans during RACE's biennial echo integration-trawl survey for walleye pollock on the EBS shelf. Biologists from NMML were able to join the RACE surveys in 1997, 1999, 2000, 2002, 2004, 2008, and 2010, providing an opportunity to describe cetacean distribution and calculate abundance over a broad area of the EBS shelf. It was possible to place observers on the entire acoustic trawl survey in only 2002, 2008, and 2010; the 2008 and 2010 surveys were part of the Bering Sea Project.
Three marine mammal observers conducted visual surveys along transect lines sampled during the NOAA walleye pollock assessment survey in June and July of 2002, 2008, and 2010. Standard line-transect survey protocols were followed, with two observers using 25× (Big Eye) reticle binoculars at port and starboard stations on the flying bridge. A third observer focused on the trackline but scanned the entire area forward of the ship by eye, or with 7×50 reticle binoculars, and recorded data using the Southwest Fisheries Science Center's software program WinCruz. Effort, environmental, and species/group data were recorded for each sighting. For abundance estimation, the study area was restricted to the U.S. side of the U.S./Russia Convention Line to make the estimates comparable across years.
Abundance estimates and sighting rates were computed for species with sufficient numbers of sightings, using multiple covariance distance sampling methods as implemented in the Mark-Recapture Distance Sampling (mrds) package for the statistical program R. Annual rates of change were estimated for each species by fitting an exponential growth model to the log of the abundance estimates.
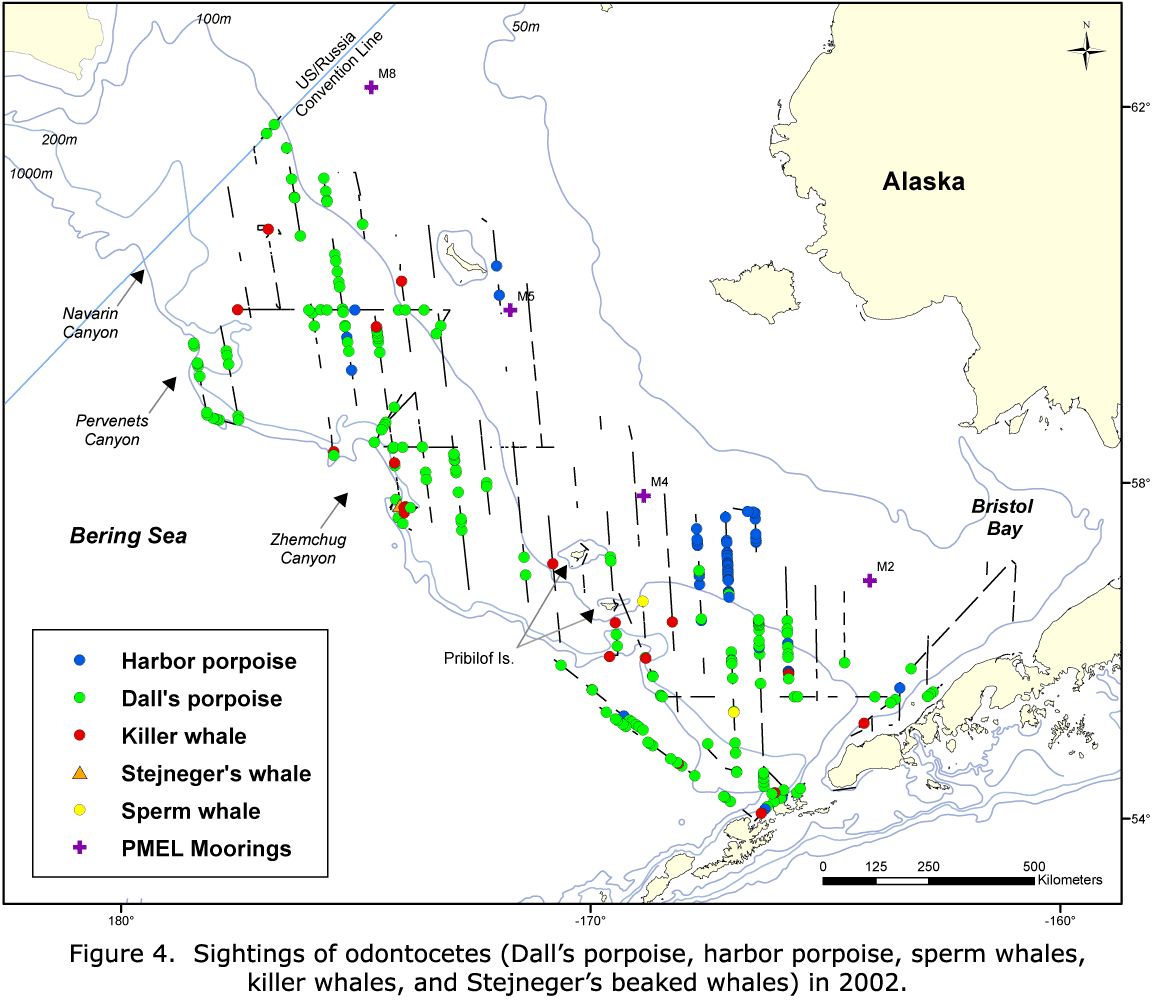
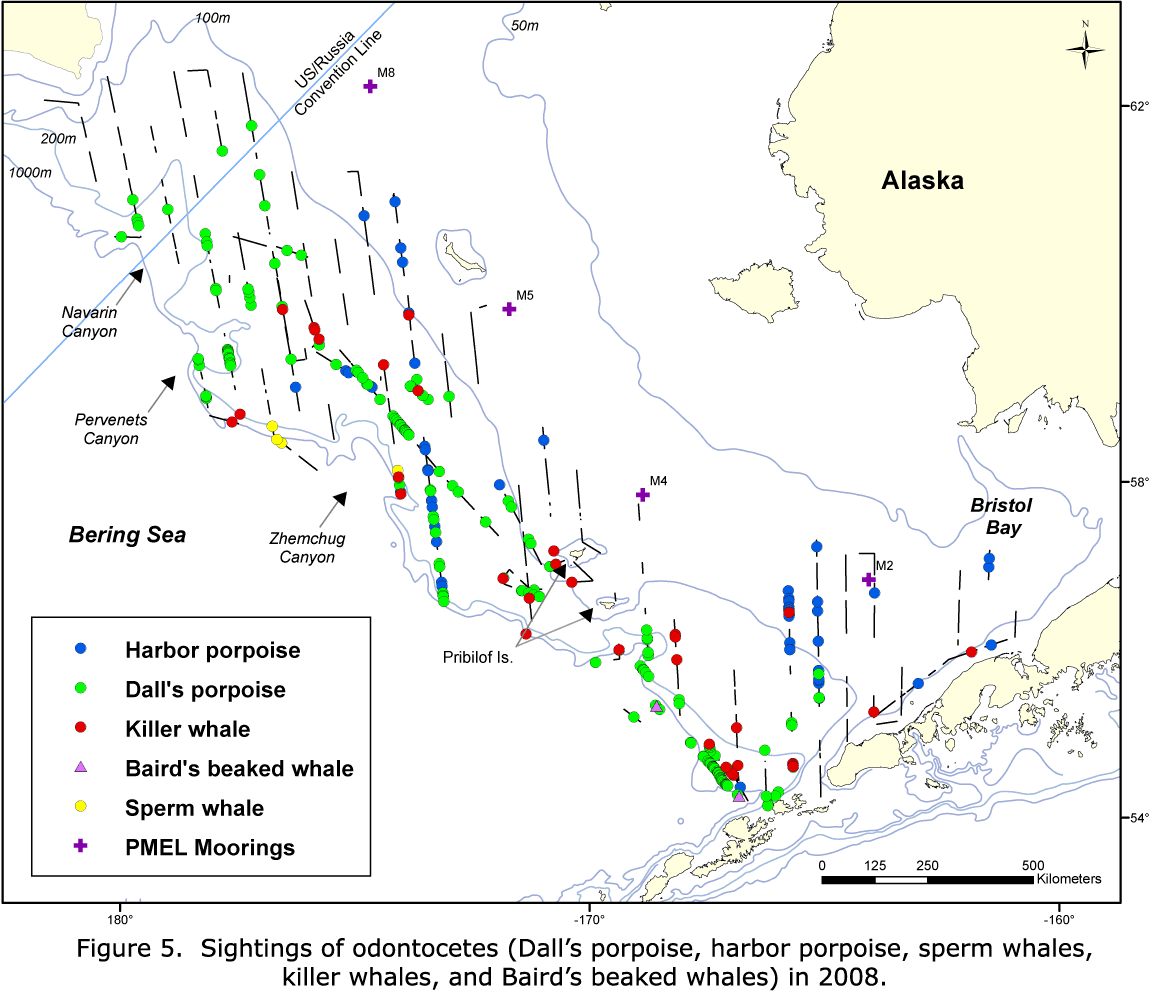
Species with a sufficient number of sightings to examine distribution and estimate abundance were humpback whales, fin whales, minke whales (Balaenoptera acutorostrata), Dall's porpoise (Phocoenoides dalli), and harbor porpoise (Phocoena phocoena). Killer whales (Orcinus orca) were widely distributed but were not identified to ecotype ("resident," "transient," or "offshore").
Less frequent sightings included sei whales (Balaenoptera borealis), gray whales (Eschrichtius robustus), sperm whales (Physeter macrocephalus), Baird's beaked whales (Berardius bairdii), Stejneger's beaked whales (Mesoplodon stejnegeri), and Pacific white-sided dolphins (Lagenorhynchus obliquidens), as well as sightings that were not identified to species.
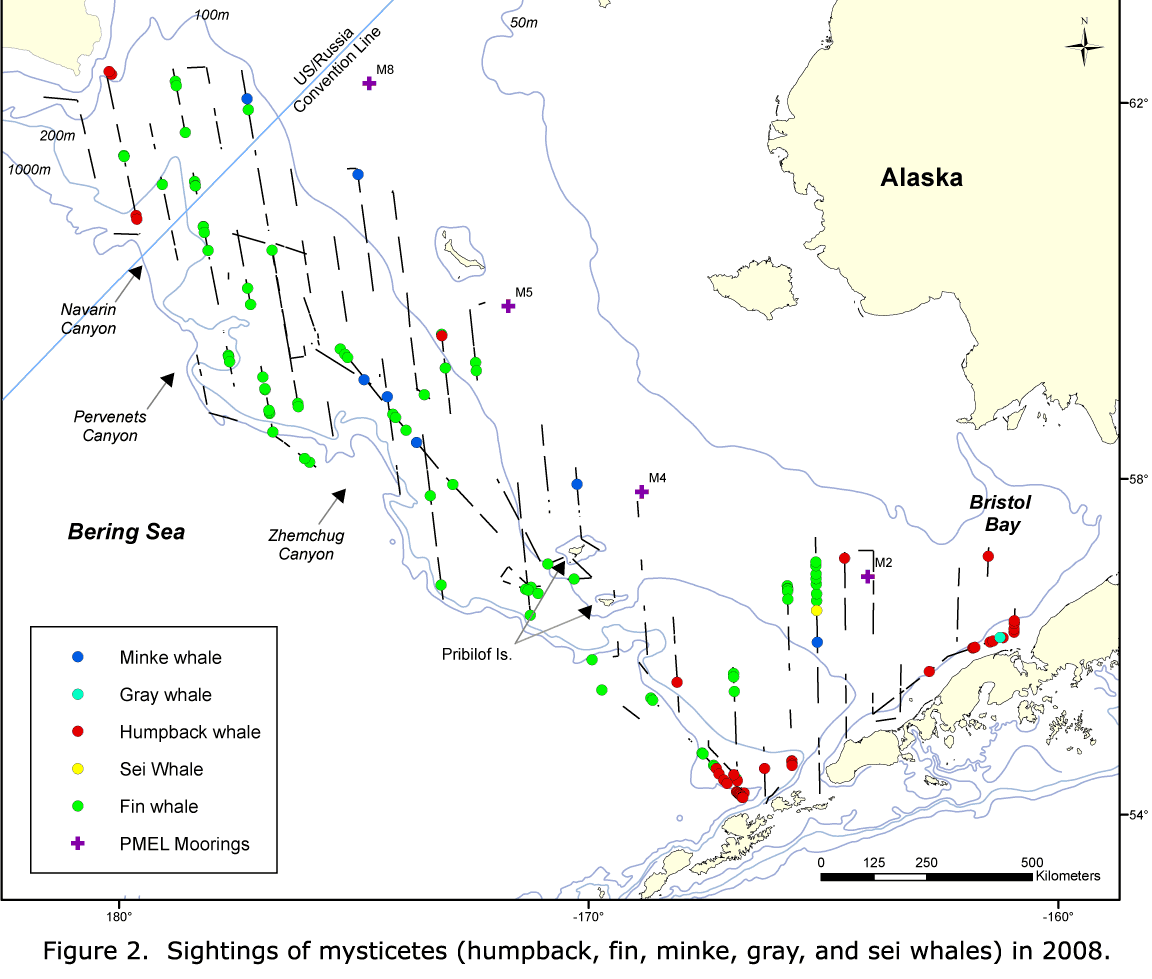
Distribution patterns for each species were similar among years, although differences in sampling effort confounded interannual comparisons, particularly east of the Pribilof Islands (Figs. 5-10). Humpback whales were consistently concentrated in coastal waters north of Unimak Pass and along the Alaska Peninsula, with only scattered sightings along the slope (Figs. 5-7).
The vessel surveys may not have captured all of the important habitat for humpback whales. Fin whales were well distributed in the outer stratum in all years but were sparse in 2002, except for a cluster of sightings around Pribilof Canyon (Figs. 5-7). There were scattered sightings in the middle domain in all years and a cluster of sightings around 57°N, 165°W in 2008. Overall, the distribution of humpback and fin whales seemed to be consistent among all 3 years, with increased numbers of sightings of these species in 2008 and 2010 compared to 2002.
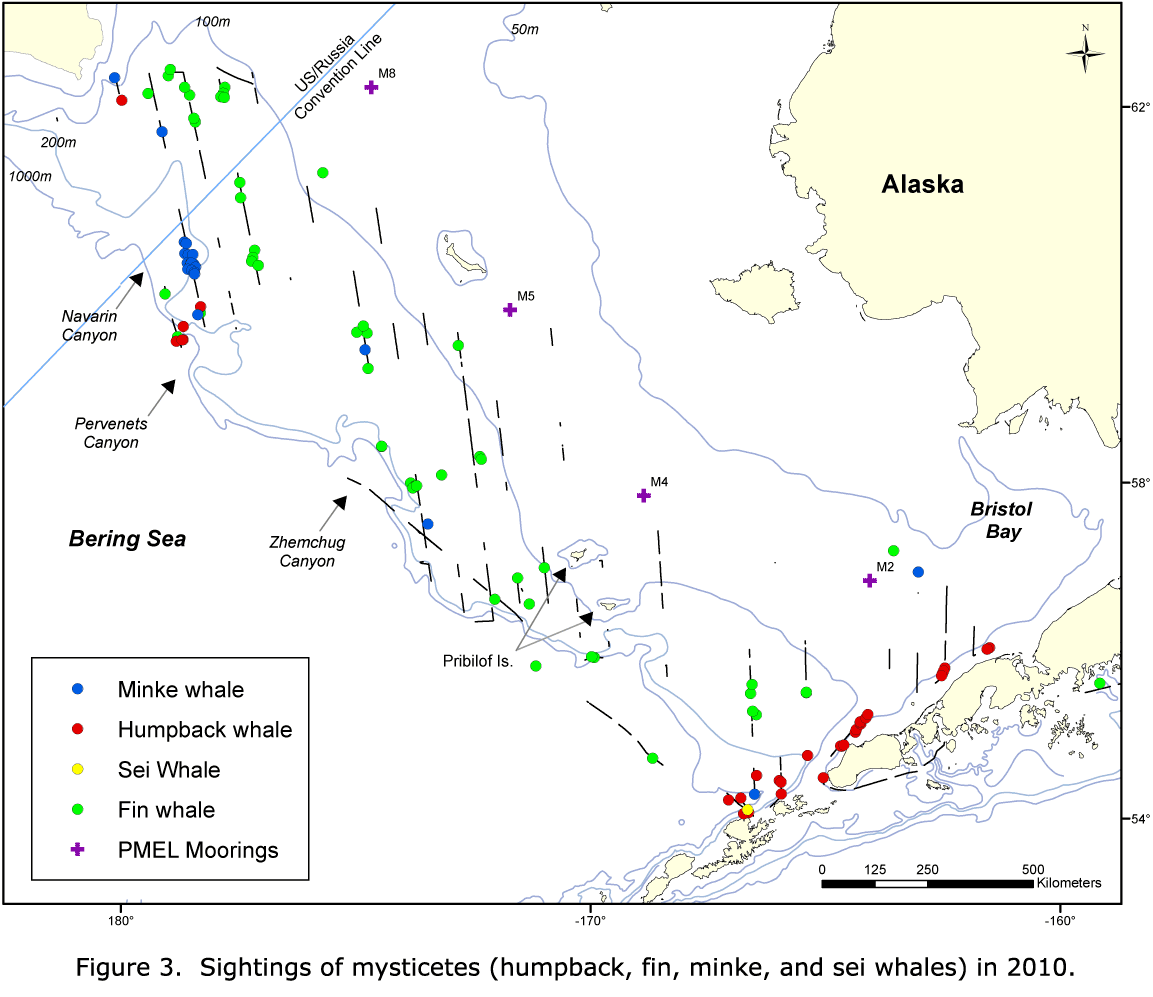
The distributions of minke whales and porpoise were more variable (Figs. 5-10). Minke whales were seen throughout the study area in all domains (Figs. 5-7). In 2002 and 2008, sightings were scattered; while in 2010, sightings were concentrated in the outer stratum in Navarin Canyon. There were roughly equal numbers of sightings in 2002 and 2010 but few in 2008.
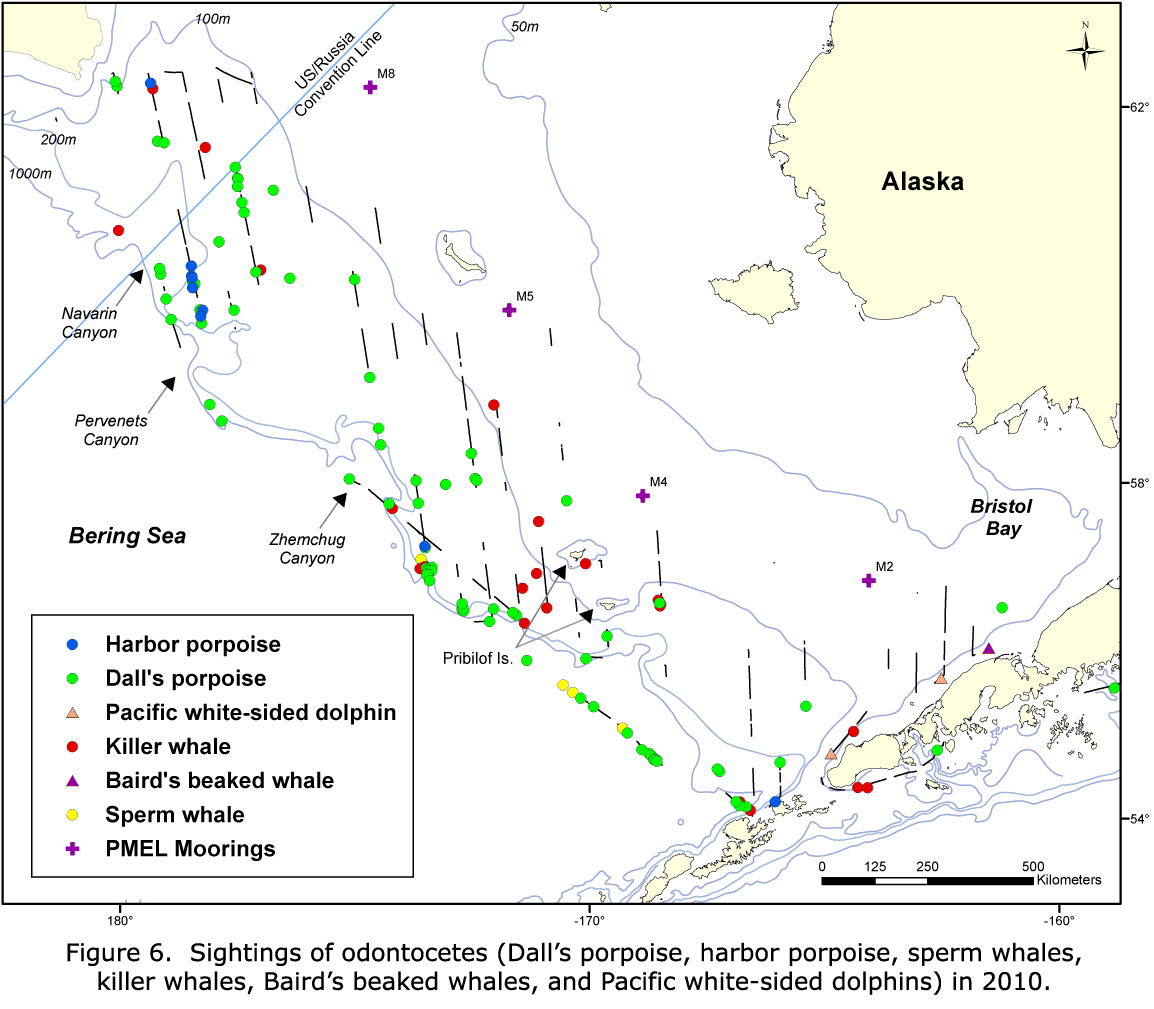
Dall's porpoise were sighted on the western edge of the middle domain and in the outer stratum in 2002; while in 2008 and 2010, all of the sightings occurred in the outer stratum (Figs. 8-10). In 2002, harbor porpoise were found in the middle domain, east of the Pribilof Islands, with scattered sightings in the outer stratum (Fig. 8). In 2008, harbor porpoise were found in the middle domain and outer stratum (Fig. 9). In 2010, there were very few harbor porpoise sightings, and they all occurred in the outer stratum, with most occurring around Pervenets and Navarin Canyons (Fig. 10).
A greater diversity of cetacean sightings in Navarin and Pervenets Canyons was observed in 2010 (Figs. 7, 10). Species not seen (minke whales and harbor porpoise) or rarely seen (humpback whales) in earlier years were conspicuously abundant. In addition, species commonly reported in previous years (Dall's porpoise and fin whales) were also present in large numbers. This region was also used by a humpback whale (satellite tagged by NMML personnel near Unalaska Island), which moved north across the Bering Sea shelf. The high abundance and diversity of cetaceans suggests that submarine canyons can be important habitats for cetaceans in the Bering Sea outer shelf and slope.
Table 1. Estimated abundance (N), coefficient of variation (CV), and 95% confidence interval (CI) for humpback whales, fin whales, minke whales, Dall's porpoise, and harbor porpoise by year.
2002
2008
2010
N
CV
95% CI
N
CV
95% CI
N
CV
95% CI
Humpback whales
231
0.63
39-1,370
436
0.45
177-1,073
672
0.80
148-3,045
Fin whales
399
0.32
210-760
1,445
0.34
738-2,831
1,155
0.38
542-2,462
Minke whales
387
0.51
146-1,030
515
0.68
146-1,818
1,936
0.73
492-7,607
Dall's porpoise
34,865
0.53
12,826-94,772
14,472
0.32
7,573-27,656
10,661
0.30
5,713-19,895
Harbor porpoise
2,031
0.46
819-5,039
3,634
0.41
1,630-8,101
728
0.66
204-2,599
Table 2. Estimated annual rate of change in
abundance (r) between 2002 and 2010 and 95%
confidence interval (CI) for humpback whales,
fin whales, minke whales, Dall's porpoise, and
harbor porpoise.
r
95% CI
Humpback whales
12.0%
-9.8 - 34.0%
Fin whales
15.8%
2.9 - 28.1%
Minke whales
15.2%
-6.2 - 37.8
Dall's porpoise
-14.8%
-29.7 - 0.1%
Harbor porpoise
-3.4%
-35.2 - 22.2%
Abundance estimates for comparable areas in 2002, 2008, and 2010 are listed in Table 1. The estimated annual rate of change in abundance of humpback, fin, and minke whales increased between 2002 and 2010 (Table 2); the increase for fin whales is statistically significant, while the increases for humpback and minke whales are not.
Given published estimated rates of increase for humpback and fin whales and what we know about the biological characteristics of humpback, fin, and minke whales, it is likely that changes in abundance in the study area are partly due to changes in distribution and not just to changes in overall population size. It is interesting to note that the Bering Sea was warmer in 2002 compared to 2008 and 2010.
In contrast, the estimated annual rate of change in abundance of Dall's and harbor porpoise decreased during this same period (Table 2); the decline for Dall's porpoise is nearly statistically significant, while the decline for harbor porpoise is not. It is unclear what could be contributing to this decrease in abundance.
There was a westward shift towards deeper water in the distribution of Dall's porpoise from 2002 to 2010 and a lack of sightings of harbor porpoise in 2010, which could indicate that the important habitat for these species has shifted outside the study area. The estimates of annual incidental mortality rate are low for observed fisheries between 2007 and 2009. However, several of the gillnet fisheries that are known to interact with these stocks are not observed.
Quantitative models are being developed to integrate data on oceanographic conditions, cetacean sightings, and prey fields. These models may shed light on whether preferred habitat shifted outside the study area in warm years for mysticetes and in cold years for odontocetes. Such studies will be important for managing cetacean species, as large positive and negative deviations from the mean trend in temperature are expected in the future.