Fisheries Behavorial Ecology Program - Newport Laboratory
Behavioral and Growth Correlates of Habitat Use in Three Co-occurring Juvenile Flatfishes
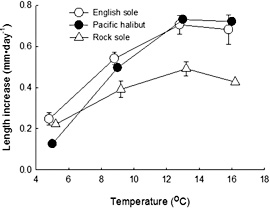
Figure 9. Growth rates of English sole, Pacific halibut, and northern rock sole in terms of mean daily length increase (+SE), at temperatures of 5°, 9°, 13° and 16°C.
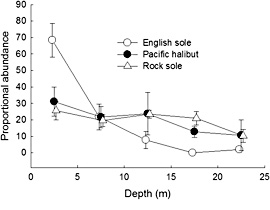
Figure 10. Mean proportional abundance (+SE) for English sole, Pacific halibut, and northern rock sole by depth at Holiday Beach, Kodiak, Alaska during July and August, 200509.
In our study we examined aspects of behavior, intrinsic growth rate, and habitat/depth distribution of the juveniles of three closely related flounder species, English sole, Pleuronectes vetulus, Pacific halibut, Hippoglossus stenolepis, and northern rock sole, Lepidopsetta polyxystra, all right-eyed flounders of the family Pleuronectidae, which co-occur in shallow water nursery embayments around Kodiak. All employ a well-developed detection-minimization strategy involving highly co-evolved morphological and behavioral adaptations.
Recent Fisheries Behavioral Ecology Program (FBEP) experiments demonstrate variation in the utilization of behavioral tactics between species. Northern rock sole (hereafter rock sole) are risk-averse; they tend to remain buried and/or prone on the bottom when not feeding, minimize movement, and do not flee unless a predator gets very close.
English sole are risk-prone; less inclined to bury, often adopt posture with their head and forward portion of the body elevated off the substrate, and more actively move about, increasing their conspicuousness to potential predators.
Pacific halibut (hereafter halibut) are risk sensitive, modifying their behavior to reflect the presence or absence of predators. In the absence of predation risk they behave like English sole. When faced with predation risk they behave more like rock sole. Standardized predation trials have demonstrated that English sole are the most vulnerable to predation, rock sole the least, with halibut intermediate between the other species.
We used these three species to test the premise that intrinsic growth and risk-taking behavior are coevolved and together influence fish distribution along a depth/predation gradient. Why? By understanding a fish's inherent behavior repertoire and scope for growth, we can better understand and characterize its essential fish habitat. First, we conducted a laboratory experiment, comparing the intrinsic growth rates of the three species. We predicted that the intrinsic growth of English sole would be greater than that of either halibut or rock sole.
Importantly, we conducted this growth experiment across a range of temperatures encompassing those encountered by fish during their nursery occupancy. As expected, rock sole grew more slowly than halibut and English sole, but this effect was temperature-dependent. Within the overall pattern of increasing growth with temperature, daily length increments (Fig. 9) were similar between species at 5°C and 9°C, but at 13°C and 16°C increments were greater for English sole and halibut than for rock sole. However, contrary to expectations, halibut growth was not lower than English sole growth at these higher temperatures.
In the second portion of this research, we utilized 5 years of field data (tows with a 2-m plumb-staff, 3-mm codend mesh) characterizing the depth distribution of the three species at one of our study sites, Holiday Beach in Kodiak, Alaska. Prior studies demonstrated a strong increase in predation pressure with depth at this site, while invertebrate biomass, and hence forage base for juvenile flatfish, increased with depth.
With a risk-prone behavioral strategy, we expected that English sole would be restricted to the shallows, where predation pressure is low. In contrast, we expected rock sole to be found deeper, where their cryptic behavior would allow them to avoid detection by predators, and because prey are more plentiful, minimize time spent foraging. We expected halibut to exhibit a depth distribution intermediate to these two. Of the three species, English sole was most clearly associated with shallow water, being more abundant at 0-5 m than at 15 25 m (Fig. 10). In contrast, both halibut and rock sole were more uniformly distributed, with no significant differences in proportional abundance across the depths examined.
Results of this study were partially consistent with our premise that risky behavior and growth are positively correlated. Rock sole are risk averse and as expected, their growth rates were the lowest of the three species. Accordingly, rock sole were more uniformly distributed across depth. With a behavioral repertoire that reduces their conspicuousness, rock sole can more successfully coexist with predators. Further, with a lower intrinsic growth rate, rock sole may adopt more of a time-minimization foraging strategy, concentrating their foraging during dusk hours, when they are less vulnerable to piscivores.
In contrast, English sole are risk prone and had a higher intrinsic growth rate. Accordingly, they are largely restricted to the shallows where predators are less abundant, and they can maximize their foraging, despite low prey densities, by feeding throughout the day. However, halibut did not match our predictions in either experiment. We believe that a broader understanding of the behavioral plasticity of juvenile halibut may help explain this apparent anomaly.
In behavioral experiments, halibut consistently demonstrate an ability to modify their behavior to match the opportunities and risks of their environment. In this study fish were reared with no predation risk. Would halibut exhibit lower growth when subjected to chronic risk? Stated another way, could perceived risk suppress growth? This perhaps is a topic for future research.
On a more applied level, our results expand knowledge of intrinsic growth rates for species of ecological and commercial importance in the north Pacific. Like other ectotherms, temperature strongly influences growth in juvenile flatfish. In the Bering Sea and Gulf of Alaska, bottom water temperature varies by season, depth, and water column structure. This variability has implications for studying the age and size structure of stocks. All three flatfish species have stocks comprised of multiple cohorts originating from broad geographic areas, in which variable temperature regimes can be expected to influence juvenile growth. While each species demonstrated a positive correlation between growth and temperature, the magnitude of the temperature response differed.
Halibut were the most temperature dependent, while rock sole were the least. Halibut growth (in terms of length) was 5.8 times greater at the highest observed rate (13°C) than at 5°C. For English sole and rock sole growth was 2.7 and 2.3 times greater, respectively. Being more influenced by temperature, halibut are probably highly dependent upon maximizing growth during the summer, when temperature in the nearshore is high. In contrast, the lower summer growth rates of rock sole may be compensated for by continued modest growth during the rest of the year.
In summary, we attempted to elucidate the relationships between anti-predator behavior, intrinsic growth rate, and depth distribution in three juvenile flatfish species that co-occur in Kodiak nurseries during the summer months. We feel that our approach of more holistically considering coevolved behavior and growth characteristics provides a useful construct for understanding juvenile fish ecology and habitat use.